
Julian Rutten, Alexander Holland and Stanislav Roudavski
Plants as Designers of Better Futures: Can Humans Let Them Lead?
ABSTRACT
This research explores the idea of plants as designers and discusses approaches that humans can use to support plant’s productive agencies. It argues that plants have unique and valuable capabilities for creating and caring for their environments. Human interventions often overlook or constrain such capabilities. In response, the article proposes to use numerical modelling to better understand plants better while challenging the anthropocentric assumptions that are common in design. It focuses on large old trees in Tasmania as examples of outstanding plant-designers that need more recognition and protection. The article also raises open questions for further research on the ethical, ecological, and aesthetic implications of vegetal design.
KEYWORDS
plant agency, large old trees, plants as designers, more-than-human care, more-than-human design
 Introduction
Introduction
Plants constitute an important part of earth’s biodiversity, which is now facing large-scale, existential threats due to human activity.1 In addition to major harms such as land use change and clearcutting, humans oppress tree lives in cities, gardens and on farmed land. They kill unwanted specimens, force trees to have shorter lives, lop off their branches, poison them, prevent their propagation, and press them into biologically uniform communities that are susceptible to parasites.2 Many humans act to protect plants but do it in ways that exclude them from decision making because dominant worldviews do not consider nonhuman beings to be intelligent, goal-oriented, and innovating organisms that act as moral agents, knowledge holders and members of multispecies polities.3 As a result, many humans resist acknowledging the roles of nonhuman beings by deploying ontological and epistemic violences when they characterise more-than-human ontologies as metaphorical or anthropomorphic. The consequence of these attitudes is a pervasive ‘plant blindness’.4 Multispecies societies become deprived of contributions that plants can make, and the resulting anthropocentric goals and approaches are corrupting the notions of justice in the society of all life and are empirically damaging at catastrophic levels.
This article aims to provide a robust account of plants as designers of better futures. We challenge three narrow assumptions about the failure of humans to protect plants, and point to distinct examples of design, creativity, and care provided by large old trees. The first assumption we challenge is that plants are not creative, decision-making agents. This assumption is based on anthropocentric presumptions that are short-sighted, exclusionary and poorly informed, as demonstrated by the repeatedly unexpected and negative consequences of human attempts at managing biotic systems.5 In contrast, we argue that plants, like other nonhuman beings, are active agents whom humans should treat with respect and appreciation. The second assumption we challenge is that plants do not contribute to design. Biomimetic designs already challenge this assumption but tend to prioritise human needs; we depart from biomimetic approaches by asking how design actions taken by nonhuman beings can contribute to all stakeholders in multispecies communities.6 The third assumption is that the inclusion of contributions by plants is impossible or impracticable.7 This is an extension of an argument that nonhuman minds or ways of being are inaccessible, especially in the case of evolutionarily distant kin, including vegetal beings. In contrast, we examine whether humans can reorient existing practices and technologies to empower nonhuman agents. We use numerical modelling as an example that demonstrates one way to incorporate design contributions of nonhuman beings such as plants.
An exploration of what humans can do to help plants help themselves as well as many other living beings requires novel methods. We draw on our expertise as researchers within a school of design8 who collaborate with scholars from fields including biology, engineering, computer science, philosophy, political science, animal studies, geography, law, anthropology and Indigenous studies. We see human traditional or scientific learning as generative of practical pathways for moral advances but are also conscious of its biases and limitations. For example, technical achievements in numerical modelling, artificial intelligence and sensing fail to benefit nonhuman beings if they progress solely in response to human interests and commercial funding.9 In contrast, this article provides an example of lidar scanning and numerical modelling as one of many ways to empower plants as political and innovating agents. Our article is situated within a developing narrative that seeks to acknowledge, understand and empower capabilities, cultures and creative contributions by nonhuman living beings in the context of interspecies or more-than-human design.
The following Section 2 of this article outlines the limitations of current approaches in a concrete design challenge. We describe the need for bird habitats, such as tree hollows, and the limitations of common knowledge systems that fail to adequately include solutions already developed by trees. Next, in Section 3, the article offers a theory of plants as empowered designers, defining agency, design, care and empowerment in ways that are deliberately non-anthropocentric. The article expands on political, philosophical and scientific developments, drawing on examples regarding Eucalyptus regnans (mountain ash) and reframing design participation to include useful contributions by nonhuman beings. Next, in Section 4, we highlight the capabilities of large old trees as design contributors, providing visual evidence of habitat structures as well as other examples of interspecies care. In Section 5, we move to examples of imaging and sensing technologies, showcasing technical workflows that can advance collaborations with plants. In Section 6, we provide an example of design actions made feasible through the proposed approach. In our conclusion, we highlight benefits and limitations of this approach as well as directions for further research and show how design in more-than-human terms can lead to pragmatic benefits and just relationships within multispecies communities.
2. Design Challenge: Future Trees in Anthropogenic Landscapes
Efforts to design artificial trees with their habitat features provides a telling example of limitations in practical design. What are the features of a good hollow for owls? Do birds prefer horizontal and dead branches? The need for evidence and meaning in observed patterns quickly reaches the boundaries of available human knowledge. Relevant knowledge sources in biology and ecology or traditional knowledge systems can provide important advice but are rarely sufficient, necessitating case-specific data collection and analysis.
One example is Barrer Hill near Canberra, Australia. This site lacks large trees and associated habitat structures. The park managers planted tens of thousands of new trees, but they will take hundreds of years to develop features that can support arboreal wildlife. In response, ecologists installed utility poles (Figure 1) that proved to be effective despite their stark structural simplicity.10 This situation is an opportunity for designs that can better replicate habitat features of biotic trees while also satisfying the need for low costs, convenient installation procedures, safety and aesthetic qualities acceptable to human communities.11
Figure 1 shows one possible design that uses an automatically generated rod-and-cable structure to simulate properties of trees.12 The advantage of such structures is their rule-based nature; human designers can use digital models to generate them in response to numerically expressed objectives and constraints. To specify such numerical targets, human designers can turn to existing successful solutions, such as branches and hollows, already developed by trees.
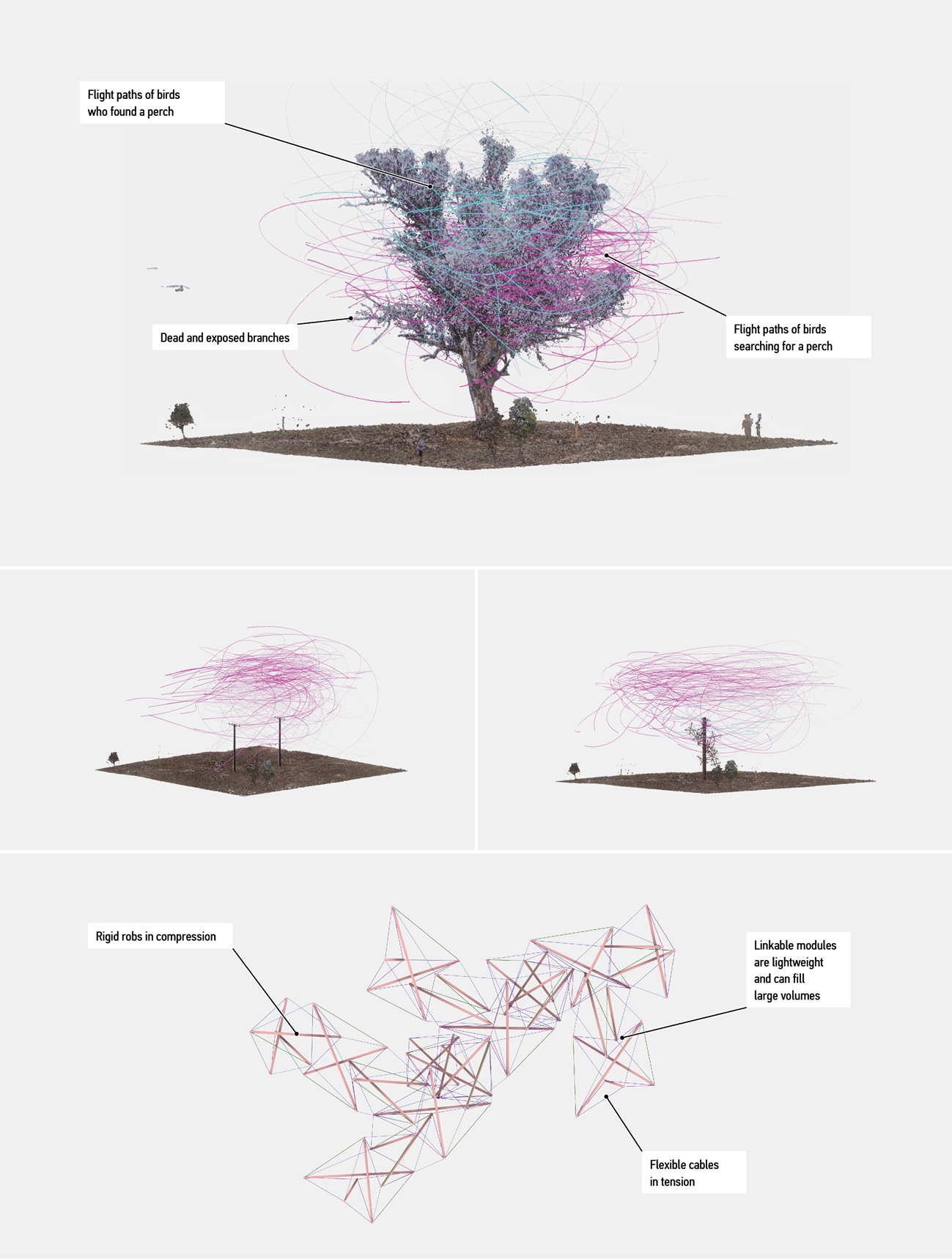
Figure 1.
Artificial habitat structures as a challenge for design. Top: a large old tree (Eucalyptus camaldulensis) provides many varied perch sites. Middle left: utility poles with a small number of artificial perch sites. Middle right: a utility pole with additional perch sites provided by a tensegrity (rod and cable) structure. Bottom: tensegrity modules. Magenta and cyan lines indicate simulated flight paths of birds searching for perches. All figures created by the authors.
3. Design Framework: Towards Plants as Empowered Designers
To consider an approach that aims to engage plants as contributors, we: 1) demonstrate that plants have agency that enables them to act as designers; 2) define the notion of design in a way that does not a priori exclude plants; 3) frame innovative and beneficial aspects of such design as forms of care that plants bestow rather than services that humans extract; and 4) consider approaches that can empower plants’ agencies in support of their lives and the lives of others.
To enable a discussion of plants as designers, we first introduce four key concepts in the list below. The subsequent sections provide evidential support for these definitions and apply them to case-study examples. In keeping with current scientific evidence and Indigenous ontologies, these definitions are deliberately non-anthropocentric; they are an attempt to include all forms of life, together with their ecosystems and communities. Nor are they anthropomorphic; on the contrary, our goal is to ‘naturalise’ human concepts and systems by demonstrating their continuity with the work of nonhuman agencies. We propose the following definitions:
Agency is a capability to act found in systems that can maintain their stability in changing conditions. Agency depends on subjective interpretations of limited information about the world provided by sensing. The notion of agency does not neatly correspond to that of an individual. Multiple agencies are always co-present and interacting.
Design is any activity that results in collaborative innovation. It is an interplay of agencies that produces novelty to cope with change. The change can be ongoing or potential, likely to occur in the future. The capability for design is significant in some circumstances but can be irrelevant or detrimental in others.
Care is a pattern of actions with shared benefits. It is a product of multiple interacting and innovating agencies. In other words, care is a possible outcome of design. Care is positive by definition; the net total of its outcomes supports life. Thus, care can serve as a criterion that captures beneficial characteristics of design.
Empowerment refers to activities that increase the ability of agents to influence political processes affecting their lives and wellbeing. In this context, empowerment leads to an ability to affect design. Often, empowerment depends on emancipation from existing systems that oppress contributions by participating agents.
3.1 Agency
To establish whether plants can design, we first need to recognise them as decision-making agents. This section relies on available evidence and theoretical frameworks that recognise directed action and meaningful interpretation as features that are necessary for and characteristic of all life.13 Here, agency is a capability to act found in systems that can maintain their stability in changing conditions. All forms of life have agency in the sense of resisting entropy.14 They process energy and matter to grow, repair and reproduce. Beyond that, an increasing body of evidence shows that all organisms, including plants, are goal-directed, intelligent, sentient, communicating and autonomous.15
For example, plants can evolve to modify their genetic composition across generations through mutation, recombination or gene flow. Genetic adaptability allows plants to cope with persistent changes in their environment, by creating new variations that may confer an advantage. Some plants have evolved resistance to herbicides, pests or diseases through gene mutation or hybridisation. Others can do this within one organism via the application of enzymes or through horizontal gene transfer.16
Plants change their behaviour, morphology and physiology in response to environmental cues, such as light, temperature, water or nutrients.17 Called phenotypic plasticity, this ability allows plants to cope with short-term or unpredictable changes in their surroundings, without altering their genetic makeup.18 For example, some plants can grow taller or produce more leaves in the shade to capture more sunlight. Other plants can change the colour or shape of their flowers to attract different pollinators.
Besides phenotypic plasticity and genetic adaptability, plants design and innovate using epigenetics, symbiosis or learning. Awareness of these abilities can be important in design because the effects of such processes might be significant but not apparent. Epigenetics refers to the changes in gene expression caused by chemical modifications or interactions with other molecules.19 For example, some plants can alter their chemical defences based on the type of herbivore that attacks them and then pass these traits to their offspring. Symbiosis refers to the close and long-term associations between different organisms. The notion of symbiosis includes mutualism, commensalism and parasitism but excludes less prolonged interactions such as predation. It is interesting here as an example of the pervasive co-evolution that results in the emergence of diverse and robust systems.20 Thus, many plants can also form mutualistic relationships with fungi, bacteria, or animals that help them obtain nutrients and water, receive protection or reproduce (Figure 3). Learning refers to the ability of a plant to modify its behaviour based on previous experience or observation. Some plants can learn from their mistakes or from other plants’ actions.21 The fuzziness and multiplicity of biological individuals make the situation even more complex.22 So does the emergence of agential actions in collectives that range from bacterial superorganisms to colonies of social insects that protect and depend on plants, as well as evolutionarily linked forest communities.
Using examples derived from the life patterns of Eucalyptus regnans (trees that we discuss in more detail later in the article and that are otherwise called mountain ash, swamp gum or stringy gum), characteristic agential interactions that often remain unknown, unnoticed and unvalued in human decision-making include:23
Self-awareness: Eucalyptus regnans do not self-fertilise promoting outcrossing, avoiding inbreeding and maintaining genetic diversity.24
Awareness of the environment: Eucalyptus regnans can sense and respond to fire by shedding large volumes of seeds into resulting ash beds in anticipation of better recruitment.25
Learning and memory: Eucalyptus regnans moderate leaf growth and accumulate nitrogen in summer to help during cooler seasons.26
Decision making: Eucalyptus regnans sprout new growth to recover after bushfires.27
Communication: Eucalyptus regnans dominate their surroundings with extensive roots that communicate and exchange nutrients with other plants and fungi.28
Niche construction: Eucalyptus regnans grow deep roots that increase water availability, reducing severe bushfires and providing habitat for other rainforest plants.29
We can conclude that plants are self-motivated, competent and innovating beings able to cope with complex communal and environmental challenges. Using multiple mechanisms that involve changes at different levels of organisation, from molecules to populations, plants are constantly evolving and experimenting with new solutions to survive and thrive in their habitats.
3.2 Design
With an outline of the agential capabilities of plants to hand, we can now provide an inclusive definition of design that does not confine design activities to professional humans. Here, our proposal is to recognise that design is any activity that results in collaborative innovation. This ‘naturalistic’ definition does not a priori confine the ability to design to one group (trained professionals, all humans or their close evolutionary relatives). Nor does it depend on the mechanisms of innovation or the attributes of the designing agent (such as the ability to plan, the possession of the theory of mind, the reliance on written texts or the use of mathematics).30
This usage of the term ‘design’ is unusual among professional designers but aligns with established interpretations in several evidence-driven domains. Relevant examples include:
– The discourse on innovation in living systems that focuses on the exploration and exploitation of ecological opportunities, phenotypic plasticity and other sources of novelty across multiple scales. Here, the term innovation encompasses the emergence of novel characteristics or processes that go beyond typical patterns of variation and selection.31
– The interpretation of processes termed niche construction within theories of Extended Evolutionary Synthesis. Niche construction is one of the common activities of all living beings.32 Processes of niche construction do not exclude human animals, their culture or their architecture.33
– The types of actions that result in habitat structures within ecosystem engineering and animal architecture.34
– The growing number of practices that self-characterise as interspecies, more-than-human, or animal-driven design, which demonstrates an increasing awareness that designing includes and depends on nonhuman beings.35
Humans often find it easier to accept that animals can act as designers under the influence of the anthropocentric bias that portrays human-like cognitive capabilities as superior. However, other forms of life also construct their niches.36 Plants specifically are remarkable designers and engineers, from planetary to microscopic scales, as shown for Eucalyptus regnans in Figure 3. In line with this reasoning, we resolve to characterise plants as designers. However, designing understood as collaborative innovation is not necessarily beneficial to whole communities and can be harmful to many stakeholders. To distinguish the beneficial characteristics of design, the next section turns to its effect that we propose to recognise as care.37
3.3 Care
Humans most frequently encounter care as a cultural, ethical or legal norm. In turn, thinking about relationships in human communities, researchers tend to define care based on their observations of human behaviours. As a result, their conceptualisations emphasise that care is a combination of human action with human emotion and, often, with human intent.38
However, many common understandings of care do not depend on human intentionality or other human attributes. For example, a broadly used definition adopted by the European Union’s European Foundation for the Improvement of Living and Working Conditions reads that care is ‘the provision of what is necessary for the health, welfare, maintenance, and protection of someone or something’.39
In keeping with this understanding, our definition suggests that care is a pattern of actions with shared benefits. Understood in this way, we avoid confining the ability to provide care to carers with privileged identities or capabilities. Instead, this definition emphasises that care is a net effect of multiple enacted relationships, rather than an outcome of intentional actions by individuals. From any local perspective, all care comes at a cost: it takes time, consumes resources, generates risks and takes away alternatives. It can involve locally negative effects including dependency, competition, parasitism and others. However, this article aims to emphasise and illustrate that over time, community-level outcomes often evolve to become positive and take the form of opportunities for diverse and persistent life.40
The benefit of this conceptualisation is in dethroning the often selfish and harmful human efforts in parallel with the acknowledgement of the supportive efforts of nonhuman others. This conceptualisation has strong empirical precedents in multiple domains. For example, traditional/Indigenous knowledge systems provide concepts such as ‘care as country’ that emphasise mutualistic support of all for all.41 Here, Country (and all that makes it up) cares for humans and other lifeforms (or sometimes it does not). If an animal, plant or human agent is attentive, they can live. If not, they will struggle. To give one interesting consequence, we agree with traditional/Indigenous views that agents do not have to be biologically alive to care. Our work shows that fallen trees provide care by linking communities or providing shelter and nutrients.42
Contemporary studies of biology and ecology increasingly concur on notions of care. For example, researchers studying constructive networks observe that all life, including trees, comes into mutualistic relationships, modifies places and assembles communities.43 This shift of emphasis from competition to facilitation and mutualism is key to seeing plants as members of care-giving communities.44 Here, we see care as the net benefit of actions in an ecological network.45
Such networks of relationships provide care when their participants can find ways to behave in patterns that support communal survival, reproduction and flourishing (Figure 4). For example, stochastic events damage limbs, which house microorganisms and over time rot into hollows to house birds or marsupials that in turn transport nutrients and seeds as they defecate. Such patterns are not guaranteed. They depend on attitudes, attributes, circumstances and luck. But the empirically observable tendency, as is the case with large old trees, is to provide and receive care.46 Such care does not need to be self-conscious, even among human beings. Attributes such as intentionality and an ability to make conscious choices are just some of many evolved tactics that can aid survival and flourishing. Brain-driven, rationalising cognitive capabilities are at the forefront of human discourse but are not necessary, universally useful or automatically beneficial.
The point of using the proposed conception of more-than-human care is to reframe the ontological position for design away from unitary entities towards networks, from objects toward processes and from humans towards heterogeneous communities of living and nonliving agents. As active and goal-directed agents, plants come into multiple interactions with other living beings. The emphasis on ‘selfish’ actions of genes or organisms tends to undervalue the supporting fabric of relationships within living communities or innovation through collaboration. In parallel with competition, activities of plants acting as ecosystem engineers enable the emergence of diverse and resistant communities.47 However, the capabilities for care vary significantly in anthropogenic environments such as managed forests, agricultural landscapes or cities.48 This is important for an understanding of the constraints on current human management and its failures to benefit from or protect contributions by plants. Recognition of care can lead to the enhancement of plants’ capacities to provide it, as we discuss in the next section.
3.4 Empowerment
Plants find themselves in the world where rapid anthropogenic changes undermine their community roles. We recognise this situation as a form of oppression that prevents better design. In the conditions of oppression, cultural appreciation, biological sciences, traditional knowledge or preservation of plants’ autonomy in wild reserves will not suffice. To contribute beneficially, it will be necessary to express and support plants’ capabilities as forms of political power that can influence decisions and redefine the future.
Plants live as striving and prospecting agents that operate subjectively in concrete situations with local information. Their interactions with other agents, including human beings, occur in constrained spaces with limited resources. These interactions are necessarily political but political representation of plants in systems such as ecodemocracy is difficult because plants cannot give consent to those who seek to represent them.49
In response, we aim to support both political representation that substitutes voices of participants and mediatic representation that reproduces and transmits their expressions. Such empowerment is necessary given the plants’ subaltern status that restricts their constitution as subjects in a polity and submits them to multiple forms of violence including structural (where plants are resources and not subjects), epistemic (where those in power presume to know better) and economic (where capital accumulation and use depend on the dispossession of plant communities). To defend their privilege, the powerful (human) agents simultaneously exclude plants and appropriate them as objects for use and exploitation.50
Political voice is an ability to engage in political representation. A qualification for such representation depends on the ability to suffer damage or endure risks emanating from policies and decisions. All those affected by such risks should have opportunities to participate in decision making.51 Existing discourse on relevant issues discusses challenges of listening to plants, ecological justice, legal standing for plants and political organisations that could include plants.52
There is a further need to move beyond acknowledgement and listening towards supporting the agential capabilities of plants to affect, innovate and benefit, but also falter and make apparently selfish decisions even when they can disadvantage or harm humans. Any meaningful redress of the current situation will require an equivalent of positive discrimination such as the compensatory principles discussed in net-positive design.53
Methods of research and design tend to privilege human agency over agencies of nonhuman beings such as plants because humans can only have partial understanding of plant actions and capabilities.54 This unavoidable limitation on human knowledge confines the scope of human leadership. As in other domains where the powerful govern the oppressed, the exclusion of stakeholder voices leads to neglect and harm. As a remedy, previous research demonstrated that co-production of knowledge can provide additional explanatory powers.55 Researchers working in the context of traditional knowledge acknowledge that a commitment to a relational ontology requires attention to co-production.56 However, others observe that being situated and attentive will not be sufficient for understanding of nonhuman lives.57
In response to these concerns, our approach is to support political and creative actions. To this end, more-than-human design can extend capabilities for beneficial innovation.58 Multiple approaches are possible, but we hypothesise that technical amplifications of human sensing, communication, analysis and action will likely play a role. To continue discussion in a concrete context, we consider lidar scanning and numerical modelling as one of many ways to empower plants as political and innovating agents.
4. Outstanding Designers: The Case of Eucalyptus regnans
Under the impact of human practices, plant species and distinct stages of plant lives are going extinct.59 For example, plants lose the ability to reproduce independently if they grow among sealed surfaces in cities or in over-fertilised, over-grazed and over-compacted agricultural landscapes. They cannot replenish the soil to provision themselves in old age or to feed others if humans remove their branches and leaf litter, as is common in managed landscapes. They cannot provide habitats to arboreal and ground-dwelling animals if they are unable to senesce and develop complex canopies that come with old age. Their abilities to reinforce the ground, create microclimates, interact with other plants and fungi or resist attacks also suffer. Some of these constrained or disappearing capabilities fall under the rubrics of ecological, functional, societal and phenotypic extinctions that are hard to recognise and address.60
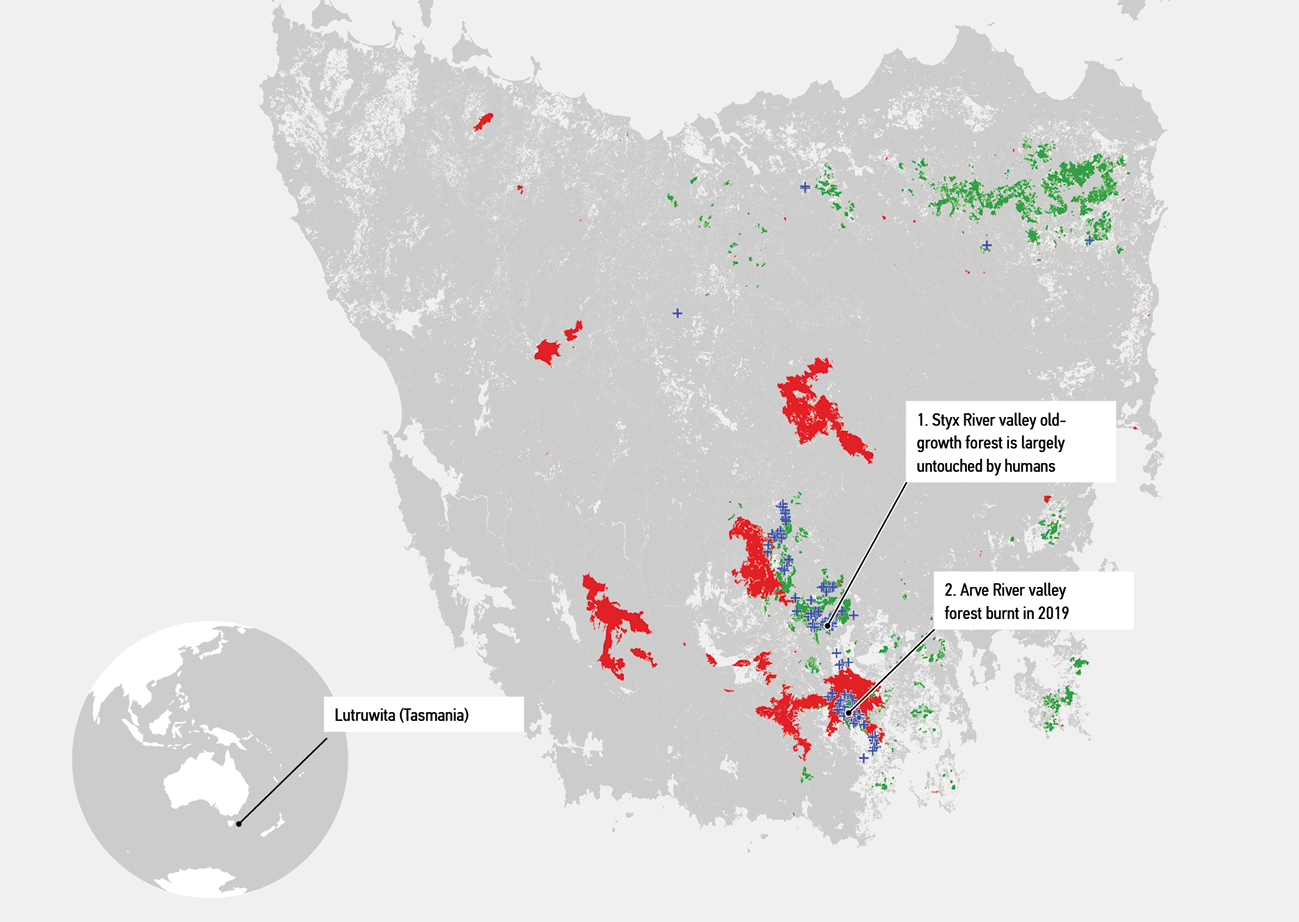
Figure 2.
Study sites. Site 1, old growth forest in the Styx Valley, and site 2, a recently burnt forest near the Arve River. Red designates the last major bushfire in 2018, while green shows Eucalyptus regnans wet sclerophyll forest. Blue crosses show trees over 70 metres or similarly old trees that have lost their tops.
To explore such challenges, we focus on large, old and rare Eucalyptus regnans trees that survive in Lutruwita (Tasmania, Australia) (Figure 2). Common in the southeast of Australia, these trees can grow to more than 100 metres and live as long as 600 years.61 Eucalyptus regnans competes for height with Sequoiadendron giganteum (giant sequoia) and is the tallest flowering plant in the world. Today, it is impossible to determine whether Eucalyptus regnans is the tallest tree on Earth because European colonists felled the biggest specimens in the 1800s without collecting the measurements. These newcomers saw Eucalyptus regnans as a source of wood for buildings, furniture and railway sleepers. Damaging human activities persist to this day. By 2019 less than one per cent of forest areas in Victoria were more than one kilometre away from disturbances such as logging and fire.62 The state of Victoria decided to end logging of native forest by 2024. In Tasmania, the logging continues. Negative impacts of climate change on forest fires further increase the already heightened vulnerability of disturbed forest communities. An important consequence is the increasing rarity of elder trees and the loss of their unique contributions. Eucalyptus regnans grow fast and may appear very tall, but most are relatively young.63 At the same time, many older trees lose height from dieback and need time without disturbances to regrow.64 We focus on large old trees because they have beneficial features that humans will do well to understand, value, support and integrate as contributions into more-than-human design (Figure 3).

Figure 3.
Design actions by a giant tree in the Styx Valley at site 1 and examples of habitat structures it provides.

Figure 4.
Examples of care from site one. Top left: bryophytes and epiphytes growing on the bark. Top right: stringy bark. Bottom left: a basal hollow. Bottom right: ground debris.
5. Promising Methods: Opportunities and Limitations of Numerical Modelling in Interspecies Design
Utilisation of creative contributions by plants in interspecies collectives will depend on broadly accepted and well-evidenced appreciation of their capabilities. Yet, plant capabilities are hard to quantify, understand and appreciate. They vary substantially and are often too complex for human analysis. Many relevant vegetal processes take hundreds or thousands of years, thus extending beyond lifespans of human projects, whole human cultures, and even the total history of human science. Plants can be small or tall or otherwise cumbersome to access and study. As a result, human knowledge about plant capabilities is lacking in detail, precision and interpretation. This lack of knowledge constrains the scope for remedial action and design innovation, limiting plants’ opportunities to contribute to design.
To illustrate one way to overcome these difficulties, we present examples of how imaging and sensing technologies, the numerical data they produce and the modelling based on these data provide ways to support design communication with nonhuman beings. The examples focus on three scales: stands of trees, individual organisms and limbs.65
We create these visualisations to obtain information about and engage with properties, capabilities and preferences of trees. To be representative such visualisations have to be products of collaborative processes that involve trees, their neighbours and meanings they exchange, which are captured as information, traced in data and recognised by algorithms.66
5.1 Data Acquisition and Preparation
A lidar device emits millions of rays of light in a spherical pattern. Each ray measures the distance by timing its return after striking a surface. Leaves block laser light, requiring multiple scans from different locations to capture all sides of a tree. Initially, these scans are independent from each other. Algorithms can align scans by looking for similar features. However, the way plants grow complicates the process. They have many self-similar features that can lead to mismatches. Branches sway in the wind and appear in different locations, creating a ghosting effect. Leaves flutter appearing as a blurry cloud rather than as distinct shapes. The height of trees also affects the data because the density of rays reduces as the light travels further away. The fine twigs, stems and leaves at height remain poorly represented. Consequently, the challenge of collecting data about trees is a difficult process that requires experience, persistence, awareness of local opportunities and constraints; in one word – craft. This craft is a dialogic endeavour that depends on previous knowledge but improves with trial and error, growing through embodied and in silico experiences with plants.
5.2 Data Analysis and Meaning Recognition
The next challenge is finding traces of meaning in the obtained information. For this to be possible, the raw data requires curation. A typical sample of 500 million points is too large for practical use with current computers. We can reduce samples by selecting points that appear sufficient for spatial analysis. Next, algorithms recognise groups of points that belong to individual trees based on the distance from their neighbours.67 This approach is quick but can miscategorise dense foliage or overlapping branches (see Figure 6). The subsequent step recognises trunk, branch and leaves by clustering points based on the similarity of their neighbours. In this case, we used a dataset of Red Tingle trees in the Southwestern part of Australia to develop a Gaussian Mixture Model measuring neighbourhood linearity, planarity, sphericality, and variation for each point. We then label clusters as tree organs.68 With a shift of perspective, we can interpret this operation as one group of trees helping humans understand another. The difference between the Red Tingle trees and Styx trees leads to some misrecognition but other measures can improve the results.
5.3 Data Visualisation and Modelling
To produce the views, the rendering system uses the sky-dome light and outlines groups of points to accentuate the depth.69 This approach can work with billions of points and supply them to the renderer on demand to avoid overwhelming the machine. It reduces the complexity of living forests to match limitations of human perception and computational capabilities of human-built machines while attempting to preserve discovered meanings. Technical capabilities and curatorial decisions by human scientists, engineers and designers can acknowledge or miss and block meanings created by the agencies of plants, opening a tricky but promising path towards mutual understanding and better collaboration.
5.3.1 Stands

Figure 5.
Complexity of old-growth forests at site one. Red shows a high vertical complexity index, yellow a medium and blue a low. Grey designates areas with no detected trees. Bottom left: the contrast between giant eucalyptus trees and the surrounding forest at site one. Bottom right: a graph that relates height, roughness and vertical complexity. Each circle represents a tree. Colours indicate vertical complexity; sizes show canopy radii. The vertical axis shows tree heights in metres and the horizontal axis plots roughness in relative units.
This example (Figure 5) visualises the measure of forest structural complexity represented by the vertical complexity index” should read “structural complexity of the forest as represented by the vertical complexity index. Structural complexity of forest stands in multiple regions relates to faunal diversity that we identify as a form of care.70 Such complexity develops in Eucalyptus regnans forests with trees older than 120 years.71 Figure 5 shows a visualisation produced by algorithms that isolate individual trees and then use aerial lidar data to estimate their complexity. For each tree, we calculate two values that relate to faunal diversity. In addition to the vertical complexity index mentioned above, we show the roughness of canopies as represented by the rumple index.72 Previous research demonstrated the value of combining multiple indices to estimate structural complexity of trees and the diagram in Figure 5 indicates the relationships between the two measures. Spatial analysis of lidar data has potential to inform the understanding of complex habitats co-designed and co-created by trees by expressing niche-constructing actions of plants in three dimensions.73 This example highlights the importance of old growth forests and helps to outline its hard-to-perceive capabilities. The resulting understanding can inform protection and management decisions, helping to resist spatial, temporal and organisational constraints of human understanding.
5.3.2 Organism

Figure 6.
A large old Eucalyptus regnans tree with its neighbourhood, trimmed to an area 50 metres wide and 50 metres deep. Colours indicate trees. Darker shades indicate leaves, lighter shades indicate wood.
The second example (Figure 6) shows a large old tree surrounded by an old-growth forest. Trees grow in response to their genetic makeup, their environment and disturbance events such as lightning, strong winds, fire or fungal attacks. As a result, each tree is unique, and shapes of large old trees can be very different.74 Numerical descriptions of tree structures in ecology and human understandings of their meaning in living communities are far from complete because such descriptions are difficult and time-consuming to produce.75 This example uses machine learning to automatically group and colour points that belong to one plant. Such categorisation can help to measure and model significant relationships in interspecies communities. Analyses of this type can help to reframe plants as designers because they focus on capabilities and innovations of trees as individuals who express the potential of their phenotypic plasticity and demonstrate the broad scope of their resilience in the face of adversity. Better understanding of these capabilities can influence assessments of health, utility, aesthetic quality, viability and other criteria that human society applies to control and often oppress the lives of trees.
5.3.3 Organs
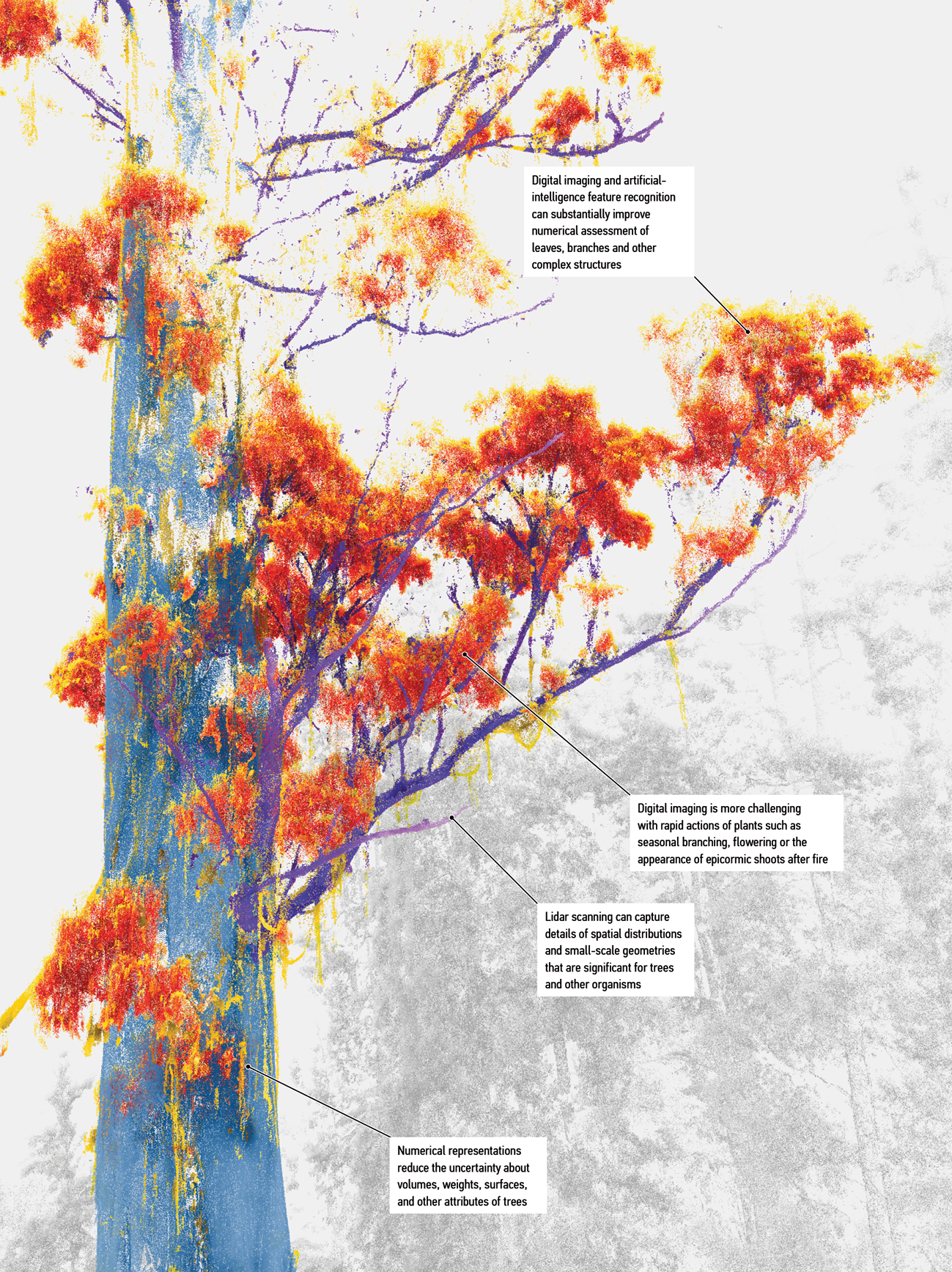
Figure 7.
Object recognition at a branch scale from site one (Figure 2). Blue, trunk; purple, branches; red, leaves; yellow, peeling bark; grey, surrounding vegetation.
This last example focuses on the structures of the canopy. Canopies support many lifeforms within buffered microclimates and complex microhabitats. To date, human sciences know them poorly but field experiments in combination with imaging can provide significant advances.76 Here, the agential perspective can emphasise capabilities that ‘canopy science’ acknowledges as forms of vegetal care. For example, bark streamers (shown as yellow in Figure 7) develop on the largest trees and are an understudied microhabitat for invertebrate taxa such as flightless tree crickets.77 In our interpretation, such structures are examples of design that can inform maintenance regimes that should preserve organic litter and provide blueprints for artificial replacements in the areas, such as many locations in inner cities, where relevant features cannot develop or persist.
***
So far, we have argued that plants are agents that design, and we illustrated some approaches that can empower such agencies. This agential framing is useful because it can provide benefits that escape other conceptualisations. Examples in the existing discourse include proposals to rely on plant mobility to resist constraints of human property, support the intellectual right of plants to promote and compensate for their contributions, frame plants as health-supporting agents in the context of ‘one health’ and limit exploitation of plants by recognising their actions as forms of labour.78
Progress in this direction will depend on persistent mutual learning. Humans can learn about chemistry, ecological engineering, climate modification, forms of mutual support and other topics. In many places, plants must find ways to live with humans and in modified landscapes. This mutual learning is a difficult challenge. Researchers studying Eucalyptus regnans forests argue for the importance of long-term projects.79 Such projects have multiple benefits but the oldest in Australia is only some 40 years in duration. By contrast, one tree can live 600 years, and three generations amount to almost 2,000.
A key benefit of lidar imaging is its ability to capture the complexity of traces produced by plant lives. For instance, lidar can identify individual plants, branches, leaves, and other parts, producing a more detailed and nuanced view of plant structures. Lidar imaging can also facilitate the interpretation of meanings expressed by plant communities. By combining lidar data from multiple perspectives and collected at different times, humans can gain a more holistic view of plants and their roles in their environment.
6. Design Response: Use of Numerical Modelling to Empower Plant Agencies
Let us now return to the design challenge introduced above. The sites and trees in it are very different from those that survive in the Styx valley, but the lessons translate. For example, our ongoing work demonstrates that it is possible to combine detailed descriptions of tree features with field observations of bird behaviours to understand the meaning of branches and other structures as they are perceived by birds.80 Numerical properties of such structures can serve as constraints or goals for generative design procedures that work with different species, structures and computational simulations including, for example, genetic algorithms, multi-agent simulations and form-finding (Figure 8).
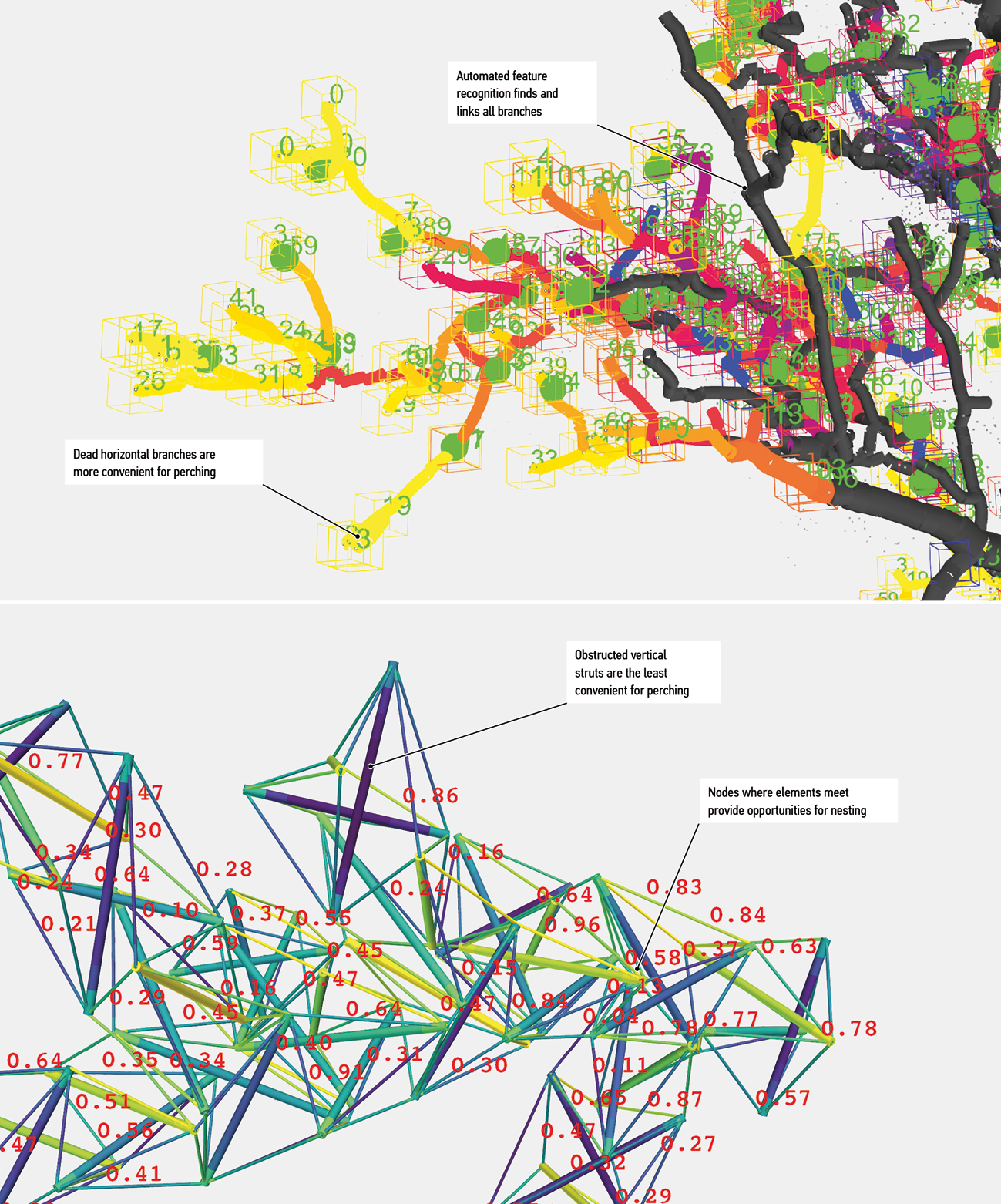
Figure 8.
Numerical analysis of living and artificial habitat structures as a basis for generative design. Numbers and colours indicate numerical estimates of suitability for perching. Top: branch segments recognised in scanned tree data. Bottom: a possible replacement design assessed against the same constraints. Yellow indicates high suitability and blue low suitability.
7. Conclusion
In this article, we asked: why do human actions fail plants and other living beings? What can humans do to help plants help themselves and many others? Our response to these questions is twofold. Firstly, we argued that even best-intentioned human practices will fall short without the inclusion of all stakeholders that can stand or benefit from their impact. This logic is similar to the convincing arguments already advanced by other emancipatory movements with their slogan of ‘nothing about us without us’.81 Secondly, we suggested that the scepticism regarding communication with and the involvement of nonhuman beings, including plants, exaggerates the barriers between forms of life on Earth. To illustrate a possible path for collaborative designing that included trees, we referred to numerical analyses of scanned data and their potential use in design. Many forms of existing learning in sciences and management already contribute to this objective but more is possible if trees, plants and other nonhuman stakeholders are supported in taking leadership roles to define new studies and experimental projects.
To support this narrative, we first described plants as decision-making agents with behaviours and actions. Secondly, we discussed plants as participants in design collectives that involve other nonhuman beings as well as humans. Thirdly, we categorised beneficial outcomes of design and as essentially supportive practices of care and indicated constraints that anthropogenic management imposes on plant capabilities. Fourthly, we demonstrated numerical imaging and modelling techniques that can strengthen plants’ capabilities but also simplify and misrepresent plant lives.
The audience for this article includes all humans who triage their attention, finances, and efforts by selecting what to study, manage, modify or support. Relevant parties include researchers in ecology and conservation science, environmental managers, designers and planners working in business or city councils, as well as experts in politics and law who oversee frameworks for the protection of biological life. Recent science and practice in combination with Indigenous and traditional worldviews challenge habitual Western opinions about agency, innovation and care in application to plants and many other beings. We hope that this article helps to advance the ongoing conversation by providing convincing justification and practical detail. It aligns with other interdisciplinary and collaborative work that includes artificial habitat-structures, manufactured replacements for disappearing old trees, smart systems to minimise environmental light pollution, urban surfaces for mosses and coastal habitats co-designed with mangroves.82
Further work in this direction is important to demonstrate that bottom-up approaches that follow living agents can interrogate and resist top-down methods of governance that entitle humans to decide what ecosystems need, which species should survive, where and how. The history of oppression across domains demonstrates that top-down paternalisms do not result in beneficial, just or sustainable outcomes and highlights the need for alternative approaches amid the increasingly acute environmental crises.
Our work shows that humans can ‘listen’ to vegetal political voices and use found meanings to recognise significant contributions that would otherwise remain unnoticed, unstudied, unappreciated and will soon disappear. This approach recognises trees not as objects, resources, or helpless patients dependent on human support, but as competent active agents, knowledge holders, innovators, and designers. This stance is different both from metaphorical appreciation of trees in human cultures as well as from biomimetic engineering that sets out to copy without compensation or acknowledgement for human use, often with harm. We hope that future research will experiment with the implications of this narrative, adjust its framing to suit the emerging evidence and formulate novel design experiments that will test and expand creative capabilities of more-than-human collectives to act – and to act with care.
ACKNOWLEDGMENTS
The authors thank many collaborators from Deep Design Lab as well as Yoav Daniel Bar-Ness, Mark Burry, Philip Gibbons, Darren Le Roux and Margaret Young for advice, practical support and the participation in the projects that informed this publication.
Author Contributions
J.R.: conceptualisation, methodology, software, validation, analysis, investigation, data curation, writing (original draft preparation, writing), review, editing, visualisation and project administration; A.H.: methodology, investigation, software, data curation, writing (review and editing, supervision); S.R.: conceptualization, methodology, validation, writing—original draft preparation, writing—review and editing, supervision, project administration, funding acquisition. All authors have read and agreed to the published version of the manuscript.
Deep Design Lab is a multidisciplinary and interspecies group dedicated to creating better worlds for all beings. Operating at the intersection of design, ecology, technology and ethics, it employs evidence-based, data-driven design experiments to develop novel theories and practices. These experiments include physical installations, computational modelling, interactive visualisation and scenario-based forecasting. The lab’s projects range from immersive storytelling about endangered plants to computational studies of tree geometries and prototypes of bioreceptive building elements. Deep Design Lab aims to develop groundbreaking solutions that foster thriving, interconnected, more-than-human communities, paving the way for more sustainable and equitable futures.
Julian Rutten is a Deep Design Lab member with qualifications in landscape architecture and mechanical engineering who conducts research at the intersection of culture, nature, and technology. His work addresses ecological challenges of urbanisation by proposing that theories of place must expand to include plants as marginalised nonhuman subjects. This research leverages the latest advances in ecology and three-dimensional imaging technologies – such as lidar, photogrammetry and computer tomography – to develop design practices that can incorporate multiple ways of knowing in support of more sustainable and just futures. Julian publishes and exhibits his work internationally and contributes to various practical environmental-conservation projects.
Email: julian.rutten@deepdesignlab.online
ORCID: 0000-0002-1427-1616
Alexander Holland is a designer and researcher at the Deep Design Lab and the University of Melbourne. Alexander studies past and future environments through collaborations with Indigenous scholars, conservation managers, designers, ecologists and data scientists. His work focuses on the political and practical implications of disappearing habitat structures, including trees. By combining methods from conservation ecology, computer science and digital design, Alexander integrates contributions from both nonhuman and human stakeholders in design processes. His research supports environmental regeneration projects and has received several awards. Alexander has presented his findings via leading scientific, artistic and architectural venues.
Email: alexander.holland@deepdesignlab.online
ORCID: 0000-0003-2922-5951
Stanislav Roudavski is an academic at the University of Melbourne and the founder of Deep Design Lab, which focuses on more-than-human design. Collaborating with scientists, engineers, creative practitioners and Indigenous scholars, Stanislav examines engagements with nonhuman agents in ethics of participation, design imagination, creative computing, digital fabrication and conservation ecology. He has participated in numerous international exhibitions and received multiple awards for excellence. Prior to his current role, Stanislav completed his PhD at the University of Cambridge and worked there on research projects. He has also taught at MIT and practised architecture in various European countries.
Email: stanislav.roudavski@deepdesignlab.online
ORCID: 0000-0003-0124-4907
1 IPBES, ‘The global assessment report on biodiversity and ecosystem services’ (Bonn: Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES) Secretariat, 2019).
2 For an example, see S. Roudavski and A. Davis, ‘Respect for old age and dignity in death: The case of urban trees’, in K. Hislop and H. Lewi (eds), What If? What Next? Speculations on History’s Futures, Proceedings of the Society of Architectural Historians Australia and New Zealand 37, (Perth: SAHANZ, 2020), pp. 638–652.
3 On plant sentience, see M. Segundo-Ortin and P. Calvo, ‘Plant sentience? Between romanticism and denial: Science’, Animal Sentience 8 (33) (2023), https://doi.org/10/gtchdm; P. Calvo and M. Segundo-Ortin, ‘Plant sentience revisited: Sifting through the thicket of perspectives’, Animal Sentience 8 (33) (2023), https://doi.org/10/gtchdj.
4 A. Amprazis and P. Papadopoulou, ‘Plant blindness: A faddish research interest or a substantive impediment to achieve sustainable development goals?’, Environmental Education Research 26 (8) (2020): 1065–1087, https://doi.org/10/ghwgn3; M. Balding and K.J.H. Williams, ‘Plant blindness and the implications for plant conservation’, Conservation Biology 30 (6) (2016): 1192–1099, https://doi.org/10/f9bpvw; C. McDonough MacKenzie, S. Kuebbing, R.S. Barak, M. Bletz, J. Dudney, B.M. McGill, M.A. Nocco, et al., ‘We do not want to “cure plant blindness” we want to grow plant love’, Plants, People, Planet 1 (3) (2019): 139–141, https://doi.org/10/gpd86f.
5 For an overview of issues, see Z. St George, The Journeys of Trees: A Story About Forests, People, and the Future (New York: W. W. Norton, 2020). On hubris, see H. Washington J. Piccolo, E. Gomez-Baggethun, H. Kopnina and H. Alberro, ‘The trouble with anthropocentric hubris, with examples from conservation’, Conservation 1 (4) (2021): 285–298, https://doi.org/10/jdjj.
6 Biomimetic designs can be as damaging as any other as is evident from the flagship example of Velcro that resulted in a huge commercial success but satisfies mostly trivial needs while generating plastic pollution and returning no benefits to plants.
7 Karl Petschke, ‘Agency without voice? A political ecology of vegetal silence’, in S. Dingli and T.N. Cooke (eds), Political Silence: Meanings, Functions and Ambiguity (New York: Routledge, 2019), pp. 129–145.
8 S. Roudavski (ed.), ‘Design for all life’, Architect Victoria 3 (2022): 32–75, https://doi.org/10/gr3wfb.
9 On the interrelationship between ways of living and states of knowledge, see S. Jasanoff (ed.), States of Knowledge: The Co-Production of Science and Social Order (London: Routledge, 2004).
10 L. Hannan et al., ‘Erecting dead trees and utility poles to offset the loss of mature trees’, Biological Conservation 236 (2019): 340–46, https://doi.org/10/ggbjtk.
11 S. Roudavski and A. Holland, ‘Tree designers and bird clients’, Landscape Architecture Australia 177 (2023): 38–43, https://doi.org/10/gthkkz. For an example of a sculptural attempt at this site, see M. Whitelaw, J. Hwang and D. Le Roux, ‘Design collaboration and exaptation in a habitat restoration project’, She Ji: The Journal of Design, Economics, and Innovation 7 (2) (2021): 223–241, https://doi.org/10/gk8x.
12 A. Holland and S. Roudavski, ‘Participatory design for multispecies cohabitation: By trees, for birds, with humans’, in S. Heitlinger, M. Foth, and R. Clarke (eds), Designing More-than-Human Smart Cities: Beyond Sustainability, Towards Cohabitation (Oxford: Oxford University Press, 2024), pp. 93–128, https://doi.org/m7z3.
13 For two overlapping approaches, see: D.M. Walsh and G. Rupik, ‘The agential perspective: Countermapping the modern synthesis’, Evolution & Development (2023): 1–18, https://doi.org/10/gsmzs3; K. Kull, ‘Biosemiotics: To know, what life knows’, Cybernetics and Human Knowing 16 (3–4) (2009): 81–88; M. Barbieri, ‘What is information?’, Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 374 (2063) (2016): 20150060, https://doi.org/10/gd6fcg.
14 See the discourse emanating from E. Schrödinger, What Is Life? The Physical Aspect of the Living Cell; with, Mind and Matter and Autobiographical Sketches (Cambridge: Cambridge University Press, 1992).
15 For some of the references on plant intelligence, see A. Trewavas, ‘Aspects of plant intelligence’, Annals of Botany 92 (1) (2003): 1–20, https://doi.org/10/bm3gx5; S. Mancuso and A. Viola, Brilliant Green: The Surprising History and Science of Plant Intelligence (Washington: Island Press, 2015); S. Mancuso, The Revolutionary Genius of Plants: A New Understanding of Plant Intelligence and Behavior, trans. V. Di Stefano (2017; repr., New York: Atria Books, 2018); Q. Hiernaux, Du comportement végétal à l’intelligence des plantes? (Versailles Cedex: Éditions Quæ, 2020).
16 P. Raimondeau et al., ‘Lateral gene transfer generates accessory genes that accumulate at different rates within a grass lineage’, New Phytologist (2023), https://doi.org/10/gsthz8.
17 T. Piersma and J.A. van Gils, The Flexible Phenotype: A Body-Centred Integration of Ecology, Physiology, and Behaviour (Oxford: Oxford University Press, 2011).
18 A. Raza et al., ‘Plant adaptation and tolerance to environmental stresses: Mechanisms and perspectives’, in M. Hasanuzzaman (ed.), Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives I: General Consequences and Plant Responses (Singapore: Springer, 2020), pp. 117–145.
19 D. Ramos-Cruz, A. N. Troyee and C. Becker, ‘Epigenetics in plant organismic interactions’, Current Opinion in Plant Biology, Epigenetics 61 (2021): 102060, https://doi.org/10/gkbncg.
20 J.N. Thompson, Interaction and Coevolution (Chicago: The University of Chicago Press, 1982); P.A. Corning, ‘A systems theory of biological evolution’, Biosystems 214 (2022): 104630, https://doi.org/10/gsf5zb.
21 For a review of perspectives and examples, see F. Baluska, M. Gagliano and G. Witzany (eds), Memory and Learning in Plants (Cham: Springer, 2018).
22 R.A. Wilson and M.J. Barker, ‘Biological individuals’, in The Stanford Encyclopedia of Philosophy, ed. Edward N. Zalta (2007; repr., Stanford: Stanford University, 2019).
23 Refer to Supplementary Material: Plant Agencies table for a more detailed listing. For the additional background of cognition beyond humans, see P. Lyon et al., ‘Reframing cognition: Getting down to biological basics’, Philosophical Transactions of the Royal Society B: Biological Sciences 376 (1820) (2021): 20190750, https://doi.org/10/ghvqh4; Segundo-Ortin and Calvo, ‘Plant sentience?’; Calvo and Segundo-Ortin, ‘Plant sentience revisited’.
24 A.R. Griffin et al., ‘Life cycle expression of inbreeding depression in Eucalyptus regnans and inter-generational stability of its mixed mating system’, Annals of Botany 124 (1) (2019): 179–187, https://doi.org/10/ghwdh3.
25 Griffin et al., ‘Life cycle expression of inbreeding depression in Eucalyptus regnans’.
26 J. Kruse et al., ‘Plasticity of leaf respiratory and photosynthetic traits in Eucalyptus grandis and E. regnans grown under variable light and nitrogen availability’, Frontiers in Forests and Global Change 3 (2020): 5, https://doi.org/10/gtg8pr.
27 S.C. Sillett et al., ‘Biomass and growth potential of Eucalyptus regnans up to 100m tall’, Forest Ecology and Management 348 (2015): 78–91, https://doi.org/10/f7fcqc.
28 A. Novoplansky, ‘What plant roots know?’, Mesenteric Organogenesis 92 (2019): 126–133, https://doi.org/10/gf4xbd.
29 D.Y.P. Tng et al., ‘Giant eucalypts – globally unique fire‐adapted rain‐forest trees?’, New Phytologist 196 (4) (2012): 1001–1014, https://doi.org/10/f4c2zk.
30 For a discussion of evolved innovations, see A. Wagner, The Origins of Evolutionary Innovations: A Theory of Transformative Change in Living Systems (Oxford: Oxford University Press, 2011).
31 For outlines of innovations in living systems, see, for example: M.E. Hochberg et al., ‘Innovation: An emerging focus from cells to societies’, Philosophical Transactions of Royal Society B 372 (1735) (2017): 20160414, https://doi.org/10/gfsp29; D.H. Erwin, ‘A conceptual framework of evolutionary novelty and innovation’, Biological Reviews (2021), https://doi.org/10/ghrk9r.
32 K. Laland, B. Matthews and M.W. Feldman, ‘An introduction to niche construction theory’, Evolutionary Ecology 30 (2) (2016): 191–202, https://doi.org/10/f8fvc9; F.J. Odling-Smee, K.N. Laland and M.W. Feldman, Niche Construction: The Neglected Process in Evolution (Princeton: Princeton University Press, 2003).
33 J.R. Kendal, J.J. Tehrani and J. Olding-Smee (eds), ‘Human niche construction’, special issue, Philosophical Transactions of the Royal Society B: Biological Sciences 366 (1566) (2011): 783–934, https://doi.org/10/bkvww3; K.N. Laland and M.J. O’Brien, ‘Cultural niche construction: An introduction’, Biological Theory 6 (3) (2011): 191–202, https://doi.org/10/gc3j7b; J. Odling-Smee and J.S. Turner, ‘Niche construction theory and human architecture’, Biological Theory 6 (3) (2011): 283–289, https://doi.org/10/gfspw4.
34 A. Hastings et al., ‘Ecosystem engineering in space and time’, Ecology Letters 10 (2) (2007): 153–164, https://doi.org/10/bxqmvd; N.J. Boogert, D.M. Paterson and K.N. Laland, ‘The implications of niche construction and ecosystem engineering for conservation biology’, BioScience 56 (7) (2006): 570–578, https://doi.org/10/c7r3gw; N.V. Coggan, M.W. Hayward and H. Gibb, ‘A global database and “state of the field” review of research into ecosystem engineering by land animals’, Journal of Animal Ecology 87 (4) (2018): 974–994, https://doi.org/10/gh48gf. On animal architecture, see M.H. Hansell, Built by Animals: The Natural History of Animal Architecture (Oxford: Oxford University Press, 2009).
35 S. Roudavski, ‘Interspecies design’, in J. Parham (ed.), Cambridge Companion to Literature and the Anthropocene (Cambridge: Cambridge University Press, 2021), pp. 147–162; W. Weisser and T. Hauck, ‘Animal-aided design: Using a species life-cycle to improve open space planning and conservation in cities and elsewhere’, BioRxiv (2017), 150359, https://doi.org/10/gfsqhg; R. Clarke et al., ‘More-than-human participation: Design for sustainable smart city futures’, Interactions 26 (3) (2019): 60–63, https://doi.org/10/gf35h5.
36 For the many examples of the inseparable intermeshing of the organism and the environment, see S.E. Sultan, Organism and Environment: Ecological Development, Niche Construction, and Adaption (New York: Oxford University Press, 2015).
37 See Supplementary Materials: Social Roles of Plants for a mapping of human and nonhuman designers and clients.
38 J.C. Tronto, Moral Boundaries: A Political Argument for an Ethic of Care (New York: Routledge, 1993).
39 Eurofund, https://www.eurofound.europa.eu/en/topic/care#:~:text=Care-may-be- globally-defined,those-with-disabilities-and-healthcare.
40 For the evolutionary dynamics of whole ecosystems, see e.g. G. Upreti, ‘Understanding ecosystem evolution and behavior’, in G. Upreti (ed.), Ecosociocentrism: The Earth First Paradigm for Sustainable Living (Cham: Springer, 2023), pp. 65–89.
41 S. Suchet-Pearson, S. Wright, K. Lloyd, L. Burarrwanga, on behalf of the Bawaka Country, ‘Caring as country: Towards an ontology of co-becoming in natural resource management’, Asia Pacific Viewpoint 54 (2) (2013): 185–197, https://doi.org/10/gfst9p; D. Ngurra et al., ‘Yanama budyari gumada: Reframing the urban to care as Darug Country in western Sydney’. Australian Geographer 50 (3) (2019): 279–293, https://doi.org/10/gjdcns.
42 J. Rutten et al., Vegetal Voices: Learning Through Making with Trees and Humans, digital video, laser scanning, digital modelling and simulation, photography, mapping, 4K video, 15min, 2023, Conference of the Society for Social Studies of Science (4S), Sea, Sky, And Land: Engaging in Solidarity in Endangered Ecologies, Honolulu and online, https://vimeo.com/883218960.
43 G. Losapio, ‘Contextualizing the ecology of plant–plant interactions and constructive networks’, AoB PLANTS 15 (4) (2023): plad035, https://doi.org/10/gtcggw.
44 For a discussion of facilitation, see T. Koffel, T. Daufresne and C.A. Klausmeier, ‘From competition to facilitation and mutualism: A general theory of the niche’, Ecological Monographs 91 (3) (2021): e01458, https://doi.org/gjwg4k.
45 On ecological networks, see G. Losapio, A. Montesinos-Navarro and H. Saiz, ‘Perspectives for ecological networks in plant ecology’, Plant Ecology & Diversity 12 (2) (2019): 87–102, https://doi.org/10/gf32k2.
46 S. Simard, Finding the Mother Tree: Discovering the Wisdom of the Forest (New York: Knopf, 2021). For roles of large old trees, see D.B. Lindenmayer, ‘Conserving large old trees as small natural features’, Biological Conservation 211 (B) (2017): 51–59, https://doi.org/10/gbn34v.
47 J.-F. Ponge, ‘Communities, ecosystem engineers, and functional domains’, Ecological Research 36 (5) (2021): 766–777, https://doi.org/10/gn5v25.
48 See Supplementary Materials: Forms of Care for forms of care by large old trees and the constraints in human-dominated environments.
49 On ecodemocrary, see J. Gray and P. Curry, ‘Ecodemocracy and political representation for non-human nature’, in H. Kopnina and H. Washington (eds), Conservation: Integrating Social and Ecological Justice (Cham: Springer, 2020), pp. 155–166. On representation, see M. Marder, Plant-Thinking: A Philosophy of Vegetal Life (New York: Columbia University Press, 2013), p. 185.
50 G. Chakravorty Spivak, ‘Can the subaltern speak?’, in G. Nelson and L. Grossberg (eds), Marxism and the Interpretation of Culture (Urbana: University of Illinois Press, 1988), pp. 271–313; M. Marder, ‘Resist like a plant! On the vegetable life of political movements’, Peace Studies Journal 5 (1) (2012): 24–32; Petschke, ‘Agency without voice?’.
51 R. Eckersley, ‘Representing nature’, in S. Alonso, J. Keane and W. Merkel (eds), The Future of Representative Democracy (Cambridge: Cambridge University Press, 2011), pp. 236–257; J. Gray and P. Curry, ‘Ecodemocracy: Helping wildlife’s right to survive’, ECOS 37 (1) (2016): 18–27.
52 A.M. Lawrence, ‘Listening to plants: Conversations between critical plant studies and vegetal geography’, Progress in Human Geography 46 (2) (2021): 629–651, https://doi.org/10/gnzjdq; A. Wienhues, Ecological Justice and the Extinction Crisis: Giving Living Beings Their Due (Bristol: Bristol University Press, 2020); C.D. Stone, ‘Should trees have standing? Toward legal rights for natural objects’, Southern California Law Review 45 (1972): 450–501; S. Mancuso and G. Conti, The Nation of Plants (2019; repr., New York: Other Press, 2021).
53 J. Birkeland, Net-Positive Design and Sustainable Urban Development (New York: Routledge, 2020).
54 V. Plumwood, ‘Nature in the active voice’, Ecological Humanities 46 (2009): 111–128; J. Atchison and L. Head, ‘Rethinking ethnobotany? A methodological reflection on human–plant research’, in M. Bastian, O. Jones, N. Moore and E. Roe (eds), Participatory Research in More-than-Human Worlds (Abingdon: Routledge, 2017), pp. 179–191.
55 Jasanoff, States of Knowledge.
56 S. Wright et al., ‘Telling stories in, through and with country: Engaging with Indigenous and more-than-human methodologies at Bawaka, NE Australia’, Journal of Cultural Geography 29 (1) (2012): 39–60, https://doi.org/10/gg4c8v.
57 Atchison and Head, ‘Rethinking ethnobotany?’.
58 S. Roudavski, ‘Multispecies cohabitation and future design’, in S. Boess, M. Cheung and R. Cain (eds), Proceedings of Design Research Society (DRS) 2020 International Conference: Synergy (London: Design Research Society, 2020), pp. 731–750, https://doi.org/10/ghj48x.
59 On large old trees, see: D.B. Lindenmayer and W.F. Laurance, ‘The ecology, distribution, conservation and management of large old trees’, Biological Reviews 92 (132) (2016): 1434–1458, https://doi.org/10/gdvpqh; M. Gilhen-Baker et al., ‘Old growth forests and large old trees as critical organisms connecting ecosystems and human health. A Review’, Environmental Chemistry Letters 20 (2022): 1529–1538, https://doi.org/10/gpv9cg. On functional extinctions in plants, see Q. Cronk, ‘Plant extinctions take time’, Science 353 (6298) (2016): 446–47, https://doi.org/10/gg879s.
60 Cronk, ‘Plant extinctions take time’.
61 A. Hay, Gum: The Story of Eucalypts and Their Champions (Sydney: NewSouth Publishing, 2021). For the background on Eucalyptus regnans, see D. Lindenmayer et al., Mountain Ash: Fire, Logging and the Future of Victoria’s Giant Forests (Clayton South: CSIRO, 2015). For the history of the struggles to preserve Tasmanian forests, including Styx, see G. Buckman, Tasmania’s Wilderness Battles: A History (Crows Nest: Allen and Unwin, 2008).
62 D. Lindenmayer et al., ‘The case for listing mountain ash forests in the central highlands of Victoria as a threatened ecological community’, Pacific Conservation Biology (2023): PC23010, https://doi.org/10/gssg7v; C. Taylor and D.B. Lindenmayer, ‘Temporal fragmentation of a critically endangered forest ecosystem’, Austral Ecology 45 (3) (2020): 340–354, https://doi.org/10/gjpxtk.
63 D. Lindenmayer and E. Bowd, ‘Critical ecological roles, structural attributes and conservation of old growth forest: Lessons from a case study of Australian mountain ash forests’, Frontiers in Forests and Global Change 5 (2022): 878570, https://doi.org/10/gr3ht8.
64 DPIPWE, ‘Giant trees and very tall forest values in the Tasmanian Wilderness World Heritage Area’, Report for the Tasmanian Wilderness World Heritage Area Natural Values Identification and Assessment Program (Hobart: Tasmanian Government, Department of Primary Industries, Parks, Water and Environment, 2018); J.L. Williams, D. Lindenmayer and B. Mifsud, ‘The largest trees in Australia’, Austral Ecology 48 (4) (2023): 653–671, https://doi.org/10/gtcq93.
65 See, Supplementary Materials: Technical Workflow.
66 On the construction of images in science, see P. Galison and C.A. Jones (eds), Picturing Science, Producing Art (Abingdon: Routledge, 1998).
67 J. Hackenberg et al., ‘SimpleTree: An efficient open source tool to build tree models from TLS clouds’, Forests 6 (11) (2015): 4245–4294, https://doi.org/10/ggb35g.
68 D. Belton, S. Moncrieff and J. Chapman, ‘Processing tree point clouds using gaussian mixture models’, ISPRS Annals of Photogrammetry, Remote Sensing and Spatial Information Sciences II-5/W2 (2013): 43–48, https://doi.org/10/gcdbx8.
69 M. Schütz, Potree: Rendering large point clouds in web browsers (Diploma Thesis, Vienna University of Technology, 2016). M. Schütz, S. Ohrhallinger and M. Wimmer, ‘Fast out‐of‐core octree generation for massive point clouds’, Computer Graphics Forum 39 (7) (2020): 155–167, https://doi.org/10/gn3gvh.
70 As implemented in the LidR package for the R programming language, also see K.Y. Van Ewijk, P.M. Treitz and N.A. Scott, ‘Characterizing forest succession in central Ontario using lidar-derived indices’, Photogrammetric Engineering and Remote Sensing 77 (3) (2011): 261–269, https://doi.org/10/gssw8q.
71 Lindenmayer and Bowd, ‘Critical ecological roles, structural attributes and conservation of old growth forest’.
72 As implemented in the LidR package for the R programming language, see also J.S. Jenness, ‘Calculating landscape surface area from digital elevation models’, Wildlife Society Bulletin 32 (3) (2004): 829–839, https://doi.org/10/fvc7bv.
73 S. Gámez and N.C. Harris, ‘Conceptualizing the 3D niche and vertical space use’, Trends in Ecology & Evolution 37 (11) (2022): 953–962, https://doi.org/10/gqkx4t.
74 F. Sterck, ‘Woody tree architecture’, in C.G.N. Turnbull (ed.), Plant Architecture and Its Manipulation (Oxford: Blackwell, 2005).
75 Y. Malhi et al., ‘New perspectives on the ecology of tree structure and tree communities through terrestrial laser scanning’, Interface Focus 8 (2) (2018): 20170052, https://doi.org/10/gf3k43.
76 C.M.P. Ozanne et al., ‘Biodiversity meets the atmosphere: A global view of forest canopies’, Science 301 (5630) (2003): 183–186, https://doi.org/10/cpftbc; A. Nakamura et al., ‘Forests and their canopies: Achievements and horizons in canopy science’, Trends in Ecology & Evolution 32 (6) (2017): 438–451, https://doi.org/10/f987zv.
77 Lindenmayer and Laurance, ‘The ecology, distribution, conservation and management of large old trees’.
78 B. Cooke and R. Lane, ‘Plant–human commoning: Navigating enclosure, neoliberal conservation, and plant Mobility in exurban landscapes’, Annals of the American Association of Geographers 108 (6) (2018): 1715–1731, https://doi.org/10/gdg5fj; D.J. Jefferson, Towards an Ecological Intellectual Property: Reconfiguring Relationships Between People and Plants in Ecuador (Abingdon: Routledge, 2020); S. Elton, ‘Relational health: Theorizing plants as health-supporting actors’, Social Science & Medicine 281 (2021): 114083, https://doi.org/10/gkbvfj; J. Palmer, ‘Putting forests to work? Enrolling vegetal labor in the socioecological fix of bioenergy resource making’, Annals of the American Association of Geographers 111 (1) (2021): 141–156, https://doi.org/10/gjhbmd.
79 Lindenmayer et al., ‘The case for listing mountain ash forests in the central highlands of Victoria as a threatened ecological community’.
80 Holland and Roudavski, ‘Participatory design for multispecies cohabitation’.
81 J.I. Charlton, Nothing About Us Without Us: Disability Oppression and Empowerment (Berkeley: University of California Press, 1998).
82 D. Parker et al., ‘A framework for computer-aided design and manufacturing of habitat structures for cavity-dependent animals’, Methods in Ecology and Evolution 13 (4) (2022): 826–841, https://doi.org/10/gpggfj; A. Holland et al., ‘Modelling and design of habitat features: Will manufactured poles replace living trees as perch sites for birds?’, Sustainability 15 (9) (2023): 7588, https://doi.org/10/gr7jqf; T. Yu and S. Roudavski, Intelligent Lighting Networks, Interactive virtual reality simulation, text, diagrams, speculative design considering interactions at urban and landscape scales, 2021, Future Implied Media Architecture Biennale, event by Amsterdam University of Applied Sciences, Utrecht University, and others, virtual; G. Tenggono, S. Sintusingha, and S. Roudavski, More-Than-Human Design for Coastal Justice: A Case Study of Mangroves in Jakarta’s Bay, 2023, Design experiments presented as a video that includes drawings, diagrams, photos, text and digital models, 4K video, 10 min, 2023, Architectural Humanities Research Association (AHRA) International Conference, Situated Ecologies of Care, Portsmouth and online.
APPENDIX: SUPPLEMENTARY MATERIALS
These Supplementary Materials provide additional evidence for the narrative in the main text. The materials include listings of: plant agencies (1), forms of design and the roles of plants (2), care (3) and an outline of the technical workflow used in the examples (4).
Plant Agencies
Table 1. Agential capabilities of nonhuman beings, including plants.
|
Agential Capability |
Example |
|
Self-awareness Recognise self. |
Plant roots distinguish between themselves, other roots of the same species, roots of different species, other organisms and minerals.A1 Empirical studies show that the ability to recognise kin and neighbours is very common.A2 In Eucalyptus regnans, an example is self-incompatibility which prevents self-fertilisation and promotes outcrossing to avoid inbreeding and maintain genetic diversity. A3 |
|
Awareness of the environment Recognise meaningful features and events in the environment. |
Plants can sense light, temperature, water, nutrients, hormones, and pathogens. They use different mechanisms to perceive these signals and translate them into biological responses.A4 For example, like many trees, Eucalyptus regnans can sense and respond to fire by shedding large volumes of seeds into resulting ash beds in anticipation of better recruitment.A5 |
|
Learning and memory Use past experiences to guide behaviour. |
For sessile organisms such as plants, cognitive processes such as learning, memory and decision-making are critical to survival and reproduction.A6 As an example, Mimosa pudica can learn not to fold its leaves in response to repeated shocks when previous exposures did not result in harm.A7 |
|
Decision making Make adaptive, discerning, flexible, anticipatory, and goal-directed decisions. |
Plants, like all organisms, act in complex environments and among conflicting signals. They routinely select one of many options and account for trade-offs.A8 For example, their roots grow away from typical gravity directed pathways to avoid toxic soils and predict dynamic nutrient patterns to maximise absorption. Plants’ epigenetic variation can function as memory, for example clonal plants interpret nutrient and light levels to forage in unknown environments.A9 |
|
Communication Exchange meaningful signals within and between biological systems. |
Plants engage in multiple forms of information processing and communication to achieve higher-level goals such as responding to pathogen attacks. A10 For example, they release and detect a wide range of volatile organic compounds to communicate with themselves, other plants, insects, fungi, microorganisms, pollinators, herbivores and their predators.A11 |
|
Niche construction Construct conditions that support life. |
Land plants alter all environments through surface remodelling and their effect on the atmosphere.A12 Plant growth, metabolism and death produce ecological niches for whole communities.A13 For example, quick growth following fires establishes eucalyptus trees whose deep roots then increase water availability, reducing severe bush fires and providing habitat for other rainforest plants.A14 |
Social Roles of Plants
To understand the roles of plants in design and outline the potential for their future engagement, we compare their current roles in design, management, and governance practices formulated by humans. Examples in this table are a simplification and many concrete projects could span multiple rows. The first row focuses on human benefits from nonhuman life and the second row explains how human management can benefit nonhuman living beings. The third row emphasises respect and protection of nonhuman lifeforms. We propose here that these existing frameworks can further benefit from a better understanding of plants’ creative agencies.
Table 2. Plants in current frameworks that govern relationships between clients and designers.
|
Designers and Clients |
Frameworks and Roles |
|
By humans for humans Design by humans to achieve human goals while minimising damage. |
Sustainable development emphasises human needs, for example through approaches that exploit forests to store carbon.A15 Planting of trees for such purposes reduces their ecological and cultural roles potentially leading to impoverished forests.A16 Similarly, nature-based solutions can aim to solve the anthropogenic heat island effect in cities by planting trees.A17 Economists frame trees’ abilities to mitigate pollution, noise and support human recreation as ecosystem services. These approaches often accept harming plants through the exposure to high temperatures and pollution but can also aim to include nonhuman interests.A18 |
|
By humans for other lifeforms Design by humans to support or restore nonhuman life. |
Some examples in this section include design for nature. This approach aims to make buildings that enhance biodiversity and create habitat niches for endangered organisms.A19 Similarly, nature positive design seeks to retrofit old designs to offset past damage and create new habitat opportunities.A20 Stepping closer to nonhuman autonomy, rewilding converts abandoned land into nature reserves with pre-human plant communities.A21 Many such approaches rely on human knowledge and judgement to improve the conditions for nonhuman lifeforms. The results can be problematic. Preference for some species can lead to the suffering of others or to the neglect of individual organisms leading to the calls for compassionate conservation and conservation welfare.A22 Respect for autonomy in bounded exclusion zones can also lead to the loss of health and death as happened in Oostvaardersplassen.A23 |
|
By all life for all life All living beings contribute to design. |
Emerging conceptions that prioritise the ‘rights of nature’ and often seek to integrate indigenous knowledge attempt to overcome human paternalism by ascribing legal and political protections to nonhuman beings. Examples include buen vivir, earth jurisprudence, animal property rights or Indigenous approaches to the care of country.A24 These important approaches frame human attitudes towards nonhuman beings and can benefit from further evidential support. Additional work to include and empower beings or relationships that operate beyond current human knowledge can further extend these approaches. |
Forms of Care
Table 3 outlines forms of care by large old trees and contrasts them with the losses that result from the curtailment of vegetal agencies.
Table 3. Forms of care and anthropogenic constraints.
|
Form of care This column includes examples of ecosystem contributions resulting from agential actions by large old trees. |
Protection of plant agency This column includes directions for future research and design that could support beneficial agencies of large old trees in the conditions of human-induced environmental change. |
|
Sustain themselves Tree roots can provide favourable environments for the growth of beneficial microorganisms, inhibit harmful nematodes and limit the growth of neighbouring plant species.A25 |
Many human activities prevent the formation and constrain the beneficial functioning of the rhizosphere. Supportive measures could include elimination of anthropogenic alterations such as hard surfaces, removal of the physical separation from conspecifics, alleviation of the stressful living conditions and restoration of important resources. These measures support trees’ ability to reproduce and maintain their young without human interventions.A26 |
|
Maintain the community Large old trees provide resources crucial to other species when alive and after their biological deaths, sometimes for hundreds or even thousands of years. This persistence supports the character of the neighbourhood and its communities.A27 |
A single old tree in an urban area can also create persistence and encourage habitation by ecologically relevant species. This ability to support the character of place also applies in other situations such as agricultural landscapes. Protective measures could include retention of trees in managed environments and a reassessment of their value.A28 |
|
Clean the environment Large old trees absorb ozone, sulphur dioxide, nitrogen dioxide and carbon dioxide while releasing oxygen through photosynthesis thus providing breathable and healthier air. They can also filter pollutants in air and water.A29 |
Trees’ ability to engage in cleaning has limits and the exposure to harmful substances can lead to stress and death. Protection of plants as agents should support trees’ right to safe environments including air, water, and soils. Protections of soils is a characteristic example where volume, compaction, contamination, structural alterations can inhibit plants’ capabilities. |
|
Regulate climate Large old trees reduce the global greenhouse effect by trapping carbon dioxide and can construct local micro- and meso- climates by providing shade and evaporating water from their leaves.A30 They further extend this effect by dropping large volumes of leaves that trap moisture and extracting underground water with their deep roots. Their impact influences rain, wind, fire, lightning, and landslides. |
Humans select plants to for protection against extreme conditions. For example, trees that act as windbreaks persist in constant stress, without an opportunity to utilise the full spectrum of their capabilities. The same is true for those deployed for their decorative value or to provide shade.A31 An extended consideration of trees’ agencies would consider these capabilities in the context of whole plant lives instead of instrumentalising them at the expense of health, wellbeing, and longevity. |
|
Supply nutrients Plants, including trees, are the primary producers in most ecosystems, converting light energy into chemical energy that is useable by other organisms. They are the main source of food for fungi, and bacteria, herbivore animals and indirectly for carnivores.A32 |
Human practices often do not allow trees to distribute nutrients though interactions with animals and fungi because anthropogenic landscapes impose accidental or deliberate isolation. Fertilisation and watering practices further reduce these abilities. Artificial light also plays a negative role by disrupting activity rhythms, increasing stress, and affecting the production of leaves, fruit and pollination networks.A33 Appropriate responses would support plants rights to live in rich communities that support a fuller spectrum of their capabilities. Here, large old trees can act as foci for communal organisations that interlink needs and entitlements of multiple lifeforms under emerging political schemas including citizenship, rights and custodianship. |
|
Support dwelling Large old trees offer protection and refuge for many animals and other organisms. They provide habitats for nesting, breeding, hiding, resting, and hunting. Some animals also use plant materials to build their own shelters, such as nests, burrows, webs and hives. For example, the hollows of large old trees can house hundreds of species of birds, mammals, reptiles, amphibians, and insects. For example, Eucalyptus regnans act as pioneer trees that can germinate after fire, establish quickly in 20-30 years, and create a microclimate that supports shade-dependent rainforest plants.A34 The canopies of large old trees contain large numbers of species many of which remain to be discovered.A35 Bark streamers are another example poorly known microhabitats. The roles of large old trees extend beyond the immediate neighbourhood and influence ecosystem processes at multiple spatial scales, including whole landscapes. |
Human practices often prevent evolved patterns of growth and senescence in way that reduce trees’ ability to contribute habitat opportunities. This happens when humans fill hollows, cut of tops to limit height, or prevent pollen, lop off branches that might fall or clean ground litter. Relevant measures can include protection of remaining old-growth forest as well as preservation, cultivation or replication of habitat features in managed landscapes.A36 |
|
Aid communication Large old trees can communicate with each other and other organisms through chemical signals, electrical impulses, sound vibrations, and physical contact. They can warn each other of pests or diseases, attract beneficial insects for pollination or protection, share resources or information with other plants or fungi and influence the behaviour or emotions of animals.A37 |
Current management practices do not take communicative capabilities of plants into account, but their consideration would substantially reframe the best case in many modified environments. For example, recent research suggests a relationship between plant volatiles and insect behaviour in the urban environment with implications for management.A38 Support for the communicative capabilities of plants can involve protection for their signalling and sensing abilities as a right to communal association. |
Technical Workflow for Empowerment
The following steps capture a typical workflow using lidar data.
Endnotes
1 Ariel Novoplansky, ‘What plant roots know?’ Mesenteric Organogenesis 92 (2019): 126–33, https://doi.org/10/gf4xbd
2 Roza D. Bilas, Amanda Bretman and Tom Bennett, ‘Friends, neighbours and enemies: An overview of the communal and social biology of plants’, Plant, Cell & Environment 44 (4) (2021): 997–1013, https://doi.org/10/gjvh2r
3 A. Rod Griffin et al., ‘Life cycle expression of inbreeding depression in Eucalyptus regnans and inter-generational stability of its mixed mating system’, Annals of Botany 124 (1) (2019): 179–87, https://doi.org/10/ghwdh3
4 Anthony Trewavas, ‘Aspects of plant intelligence’ Annals of Botany 92 (1) (2003): 1–20, https://doi.org/10/bm3gx5; Paco Calvo et al., ‘Plants are intelligent, here’s how’, Annals of Botany 125 (1) (2020): 11–28, https://doi.org/10/ggh883
5 Griffin et al., ‘Life cycle expression of inbreeding depression in Eucalyptus regnans’.
6 Frantisek Baluska, Monica Gagliaano and Guenther Witzany (eds), Memory and Learning in Plants (Cham: Springer, 2018).
7 Monica Gagliano et al., ‘Experience teaches plants to learn faster and forget slower in environments where it matters’, Oecologia 1 (2014): 63–72, https://doi.org/10/f5zqgm
8 Richard Karban and John L. Orrock, ‘A judgment and decision-making model for plant behavior’, Ecology 99 (9) (2018): 1909–19, https://doi.org/10/gdzt75; Jonny Lee, Miguel Segundo-Ortin and Paco Calvo, ‘Decision making in plants: A rooted perspective’, Plants 12 (9) (2023): 1799, https://doi.org/10/gtfj79
9 Miguel Segundo-Ortin and Paco Calvo, ‘Consciousness and cognition in plants’, WIREs Cognitive Science 13 (2) (2022): e1578, https://doi.org/10/gmwqjh
10 Ian T. Baldwin, ‘Plant volatiles’, Current Biology 20 (9) (2010): R392–97, https://doi.org/10/c6hxcm; Gen-Ichiro Arimura and Ian S. Pearse, ‘From the lab bench to the forest: Ecology and defence mechanisms of volatile-mediated “talking trees”, in Guillaume Bécard (ed.), How Plants Communicate with Their Biotic Environment (Academic Press, 2017), pp. 3–17.
11 Velemir Ninkovic, Dimitrije Markovic and Merlin Rensing, ‘Plant volatiles as cues and signals in plant communication’, Plant, Cell & Environment 44 (4) (2021): 1030–43, https://doi.org/10/gkczpr
12 Tais W. Dahl and Susanne K.M. Arens, ‘The impacts of land plant evolution on Earth’s climate and oxygenation state – an interdisciplinary review’, Chemical Geology 547 (2020): 119665, https://doi.org/10/ghn477
13 Xiaoli Dong, ‘Evolution of plant niche construction traits in biogeomorphic landscapes’, The American Naturalist 199 (6) (2022): 758–75, https://doi.org/10/grztxn
14 David Y.P. Tng et al., ‘Giant eucalypts – globally unique fire‐adapted rain‐forest trees?’ New Phytologist 196 (4) (2012): 1001–14, https://doi.org/10/f4c2zk
15 Wytze van der Gaast, Richard Sikkema and Moriz Vohrer, ‘The contribution of forest carbon credit projects to addressing the climate change challenge’, Climate Policy 18 (1) (2018): 42–48, https://doi.org/10/gm5s6t
16 Xiaojuan Liu et al., ‘Tree species richness increases ecosystem carbon storage in subtropical forests’, Proceedings of the Royal Society B: Biological Sciences 285 (1885) (2018): 20181240, https://doi.org/10/gft78w
17 Alexander Thomas Hayes et al., ‘Nature-based solutions (NBSs) to mitigate Urban Heat Island (UHI) effects in Canadian cities’, Buildings 12 (7) (2022): 925, https://doi.org/10/gssxg9
18 Carsten Herrmann-Pillath et al., ‘Nature-based solutions as more-than-human art: Co-evolutionary and co-creative design approaches’, Nature-Based Solutions 4 (2023): 100081, https://doi.org/10/gsh3fk; Cecily Maller, ‘Re-orienting nature-based solutions with more-than-human thinking’, Cities 113 (2021): 103155, https://doi.org/10/gk94tz
19 Chiara Catalano et al., ‘Smart sustainable cities of the new millennium: Towards design for Nature’, Circular Economy and Sustainability 1 (3) (2021): 1053–86, https://doi.org/10/gqc486
20 Janis Birkeland, ‘Nature positive: Interrogating sustainable design frameworks for their potential to deliver eco-positive outcomes’, Urban Science 6 (2) (2022): 35, https://doi.org/10/gqgx77
21 Andrea Perino et al., ‘Rewilding complex ecosystems’, Science 364 (6438) (2019): eaav5570, https://doi.org/10/gfzxfg
22 Ngaio J. Beausoleil, ‘I am a compassionate conservation welfare scientist: Considering the theoretical and practical differences between compassionate conservation and conservation welfare’, Animals 10 (2) (2020): 257, https://doi.org/10/ggvkrh
23 Helen Kopnina, Simon R.B. Leadbeater, and Paul Cryer, ‘Learning to rewild: Examining the failed case of the Dutch “new wilderness” Oostvaardersplassen’, International Journal of Wilderness 25 (3) (2019); Jamie Lorimer and Clemens Driessen, ‘Wild experiments at the Oostvaardersplassen: Rethinking environmentalism in the Anthropocene’, Transactions of the Institute of British Geographers 39 (2) (2014): 169–81, https://doi.org/10/f54s4b
24 Eduardo Gudynas, ‘Buen vivir: Today’s tomorrow’, Development 54 (4) (2011): 441–47, https://doi.org/10/fxvz2f; Martha Chaves et al., ‘Radical ruralities in practice: Negotiating buen vivir in a Colombian network of sustainability’, Journal of Rural Studies 59 (2018): 153–62, https://doi.org/10/ggt326; Peter D. Burdon, ‘A theory of Earth jurisprudence’, Australian Journal of Legal Philosophy 37 (2012): 28–60; Deborah Bird Rose, Nourishing Terrains: Australian Aboriginal Views of Landscape and Wilderness (Canberra: Australian Heritage Commission, 1996); Karen Bradshaw, ‘Animal property rights’, University of Colorado Law Review 89 (3) (2018): 809–62, https://doi.org/10/gfsp3n
25 Doris Vetterlein et al., ‘Rhizosphere spatiotemporal organization: A key to rhizosphere functions’, Frontiers in Agronomy 2 (2020): 00008, https://doi.org/10/gssxsf; Zoe G. Cardon and Julie Lynn Whitbeck (eds), The Rhizosphere: An Ecological Perspective (Amsterdam: Elsevier Academic Press, 2007)
26 Chi Yung Jim, ‘Urban heritage trees: Natural-cultural significance informing management and conservation’, in Puay Yok Tan and Chi Yung Jim, Greening Cities: Forms and Functions, Advances in 21st Century Human Settlements (Singapore: Springer, 2017), pp. 279–305.
27 Gianluca Piovesan and Franco Biondi, ‘On tree longevity’, New Phytologist 231 (4) (2021): 1318–37, https://doi.org/10/gnzfqd
28 Darren S. Le Roux et al., ‘The future of large old trees in urban landscapes’, PLOS ONE 9 (6) (2014): e99403, https://doi.org/10/f6dg7p
29 Peter Thomas, Trees: Their Natural History, 2nd ed. (2000; repr., Cambridge: Cambridge University Press, 2014).
30 David Lindenmayer et al., ‘Stand age related differences in forest microclimate’, Forest Ecology and Management 510 (2022): 120101, https://doi.org/10/gtfp4t
31 Tianyu Hu et al., ‘Quantifying the shape of urban street trees and evaluating its influence on their aesthetic functions based on mobile lidar data’, ISPRS Journal of Photogrammetry and Remote Sensing 184 (2022): 203–14, https://doi.org/10/gn7n6g
32 Pooja Nisha et al., ‘Biomass from terrestrial environments’, in Sabu Thomas et al., Handbook of Biomass (Singapore: Springer, 2023), pp. 1–27.
33 Dirk Sanders et al., ‘How artificial light at night may rewire ecological networks: Concepts and models’, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 378 (1892) (2023): 20220368, https://doi.org/10/gs247k
34 Tng et al., ‘Giant Eucalypts’.
35 Claire M.P. Ozanne et al., ‘Biodiversity meets the atmosphere: A global view of forest canopies’, Science 301 (5630) (2003): 183–86, https://doi.org/10/cpftbc
36 Le Roux et al., ‘The future of large old trees in urban landscapes’.
37 Alana R.O. Chin, Janneke Hille Ris Lambers and Jerry F. Franklin, ‘Context matters: Natural tree mortality can lead to neighbor growth release or suppression’, Forest Ecology and Management 529 (2023): 120735, https://doi.org/10/gtfpmw
38 Noboru Masui et al., ‘Ozone disrupts the communication between plants and insects in urban and suburban areas: An updated insight on plant volatiles’, Journal of Forestry Research 32 (4) (2021): 1337–49, https://doi.org/10/gjm4g6
39 David Belton, Simon Moncrieff and Jane Chapman, ‘Processing tree point clouds using Gaussian mixture models’, ISPRS Annals of Photogrammetry, Remote Sensing and Spatial Information Sciences II-5/W2 (2013): 43–48, https://doi.org/10/gcdbx8
40 Jan Hackenberg et al., ‘SimpleTree: An efficient open source tool to build tree models from TLS clouds’, Forests 6 (11) (2015): 4245–94, https://doi.org/10/ggb35g
41 Mark Harrower and Cynthia A. Brewer, ‘ColorBrewer.Org: An online tool for selecting colour schemes for maps’, The Cartographic Journal 40 (1) (2003): 27–37, https://doi.org/10/ch4c57; Daniel Giradeau-Montaut, ‘CloudCompare’, 2018: http://www.cloudcompare.org/.
42 Jeff W. Atkins et al., ‘Quantifying vegetation and canopy structural complexity from terrestrial lidar data using the Forestr R package’, Methods in Ecology and Evolution 9 (10) (2018): 2057–66, https://doi.org/10/gff6wv